
New paper: N2pc versus TELAS (target-elicited lateralized alpha suppression)
/Bacigalupo, F., & Luck, S. J. (in press). Lateralized suppression of alpha-band EEG activity as a mechanism of target processing. The Journal of Neuroscience. https://doi.org/10.1523/JNEUROSCI.0183-18.2018
Since the classic study of Worden et al. (2000), we have known directing attention to the location of an upcoming target leads to a suppression of alpha-band EEG activity over the contralateral hemisphere. This is usually thought to reflect a preparatory process that increases cortical excitability in the hemisphere that will eventually process the upcoming target (or decreases excitability in the opposite hemisphere). This can be contrasted with the N2pc component, which reflects the focusing of attention onto a currently visible target (reviewed by Luck, 2012). But do these different neural signals actually reflect similar underlying attentional mechanisms? The answer in a new study by Felix Bacigalupo (now on the faculty at Pontificia Universidad Catolica de Chile) appears to be both “yes” (the N2pc component and lateralized alpha suppression can both be triggered by a target, and they are both influenced by some of the same experimental manipulations) and “no” (they have different time courses and are influenced differently by other manipulations).
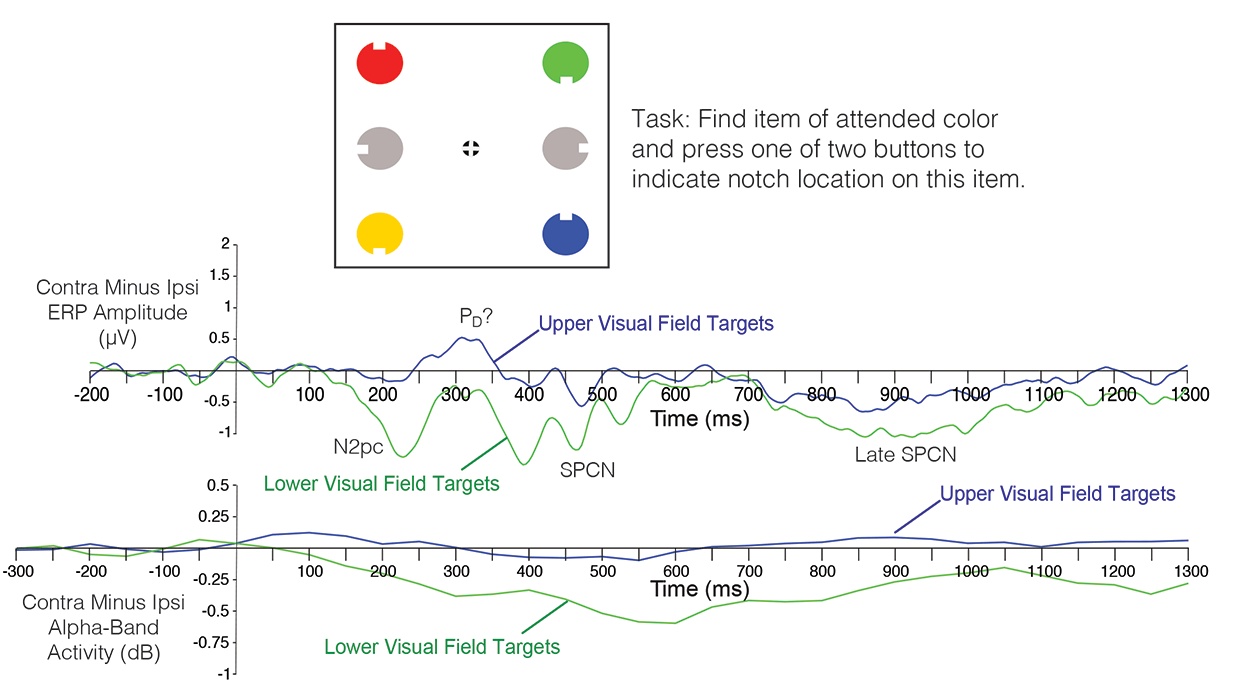
The study involved two experiments that we were designed to determine whether (a) lateralized alpha suppression would be triggered by a target in a visual search array, and (b) whether this effect could be experimentally dissociated from the N2pc component. The first experiment (shown in the figure below) used a fairly typical N2pc design. Subjects searched for an item of a specific color for a given block of trials. The target color appeared (unpredictably) at one of four locations. Previous research has shown that the N2pc component is primarily present for targets in the lower visual field, and we replicated this result (see ERP waveforms below). We also found that, although alpha-band activity was suppressed over both hemispheres following target presentation, this suppression was greater over the hemisphere contralateral to the target. Remarkably, like the N2pc component, the target-elicited lateralized alpha suppression (TELAS) occurred primarily for targets in the lower visual field. However, the time course of the TELAS was quite different from that of the N2pc. The scalp distribution of the TELAS also appeared to be more posterior than that of the N2pc component (although this was not formally compared).
The second experiment included a crowding manipulation, following up on a previous study in which the N2pc component was found to be largest when flanked by distractors that are at the edge of the crowding range, with a smaller N2pc when the distractors are so close that they prevent perception of the target shape (Bacigalupo & Luck, 2015). We replicated the previous result, but we saw a different pattern with the lateralized alpha suppression: The TELAS effect tended to increase progressively as the flanker distance decreased, with the largest magnitude for the most crowded displays. Thus, the TELAS effect appears to be related to difficulty or effort, whereas the N2pc component appears to be related to whether or not the target is successfully selected.
The bottom line is that visual search targets trigger both an N2pc component and a contralateral suppression of alpha-band EEG oscillations, especially when the targets are in the lower visual field, but the N2pc component and the TELAS effect can also be dissociated, reflecting different mechanisms of attention.
These results are also relevant for the question of whether lateralized alpha effects reflect an increase in alpha in the nontarget hemisphere to suppress information that would otherwise be processed by that hemisphere or, instead, a decrease in alpha in the target hemisphere to enhance the processing of target information. If the TELAS effect reflected processes related to distractors in the hemifield opposite to the target, then we would not expect it to be related to whether the target was in the upper or lower field or whether flankers were near the target item. Thus, the present results are consistent with a role of alpha suppression in increasing the processing of information from the target itself (see also a recent review paper by Josh Foster and Ed Awh).
One interesting side finding: The contralateral positivity that often follows the N2pc component (similar to a Pd component) was clearly present for the upper-field targets. It was difficult to know the amplitude of this component for the lower-field targets given the overlapping N2pc and SPCN components, but the upper-field targets clearly elicited a strong contralateral positivity with little or no N2pc. This provides an interesting dissociation between the post-N2pc contralateral positivity and the N2pc component.
