New Book: Applied ERP Data Analysis
/I’m excited to announce my new book, Applied ERP Data Analysis. It’s available online FOR FREE on the LibreTexts open source textbook platform. You can cite it as: Luck, S. J. (2022). Applied Event-Related Potential Data Analysis. LibreTexts. https://doi.org/10.18115/D5QG92
The book is designed to be read online, but LibreTexts has a tool for creating a PDF. You can then print the PDF if you prefer to read on paper.
I’ve aimed the book at beginning and intermediate ERP researchers. I assume that you already know the basic concepts behind ERPs, which you can learn from my free online Intro to ERPs course (which takes 3-4 hours to complete).
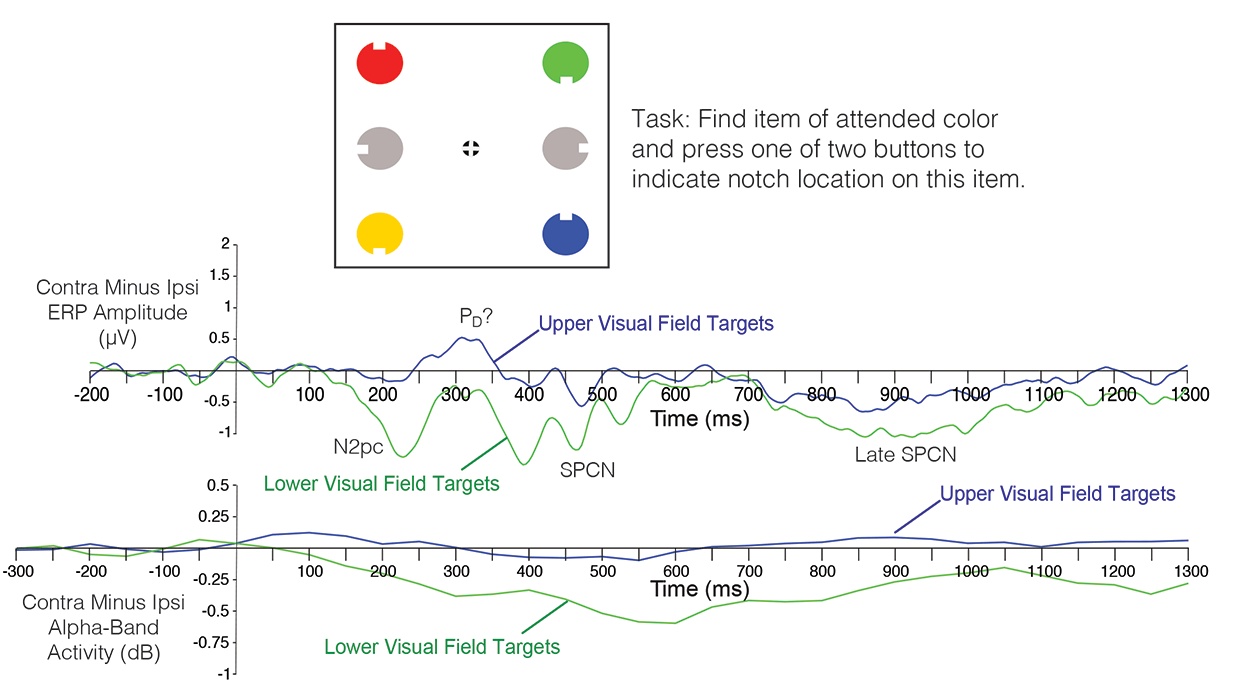
Whereas my previous book focuses on conceptual issues, the new book focuses on how to implement these concepts with real data. Most of the book consists of exercises in which you process data from the ERP CORE, a set of six ERP paradigms that yield seven different components (P3b, N400, MMN, N2pc, N170, ERN, LRP). Learn by doing!
With real data, you must deal with all kinds of weird problems and make many decisions. The book will teach you principled approaches to solving these problems and making optimal decisions.
Side note: my approach in this book was inspired by Mike X Cohen’s excellent book, Analyzing Neural Time Series Data: Theory and Practice.
You will analyze the data using EEGLAB and ERPLAB, which are free open source Matlab toolboxes. Make sure to download version 9 of ERPLAB. (You may need to buy Matlab, but many institutions provide free or discounted licenses for students.) Although you will learn a lot about these specific software packages, the exercises and accompanying text are designed to teach broader concepts that will translate to any software package (and any ERP paradigm). The logic is much more important than the software!
One key element of the approach, however, is currently ERPLAB-specific. Specifically, the book frequently asks whether a given choice increases or decreases the data quality of the averaged ERPs, as quantified with the Standardized Measurement Error (SME). If this approach makes sense to you, but you prefer a different analysis package, you should encourage the developers of that package to implement SME. All our code is open source, so translating it to a different package should be straightforward. If enough people ask, they will listen!
The book also contains a chapter on scripting, plus tons of example scripts. You don’t have to write scripts for the other chapters. But learning some simple scripting will make you more productive and increase the quality, innovation, and reproducibility of your research.
I made the book free and open source so that I could give something back to the ERP community, which has given me so much over the years. But I’ve discovered two downsides to making the book free. First, there was no copy editor, so there are probably tons of typos and other errors. Please shoot me an email if you find an error. (But I can’t realistically provide tech support if you have trouble with the software.) Second, there is no marketing budget, so please spread the word to friends, colleagues, students, and billionaire philanthropists.
This book was also designed for use in undergrad and grad courses. The LibreTexts platform makes it easy for you to create a customized version of the book. You can reorder or delete sections or whole chapters. And you can add new sections or edit any of the existing text. It’s published with a CC-BY license, so you can do anything you want with it as long as you provide an attribution to the original source. And if you don’t like some of the recommendations I make in the book, you can just change it to say whatever you like! For example, you can add a chapter titled “Why Steve Luck is wrong about filtering.”
If you are a PI: the combination of the online course, this book, and the resources provided by PURSUE give you a great way to get new students started in the lab. I’m hoping this makes it easier for faculty to get more undergrads involved in ERP research.