New papers on the hyperfocusing hypothesis of cognitive dysfunction in schizophrenia
/Luck, S. J., Hahn, B., Leonard, C. J., & Gold, J. M. (2019). The hyperfocusing hypothesis: A new account of cognitive dysfunction in schizophrenia. Schizophrenia Bulletin, 45, 991–1000. https://doi.org/10.1093/schbul/sbz063
Luck, S. J., Leonard, C. J., Hahn, B., & Gold, J. M. (2019). Is selective attention impaired in schizophrenia? Schizophrenia Bulletin, 45, 1001–1011. https://doi.org/10.1093/schbul/sbz045

SCHIZOPHRENIA BY ANNDEEF, http://fav.me/d4ggpqk, (Creative Commons License)
The most distinctive symptoms of schizophrenia are hallucinations, delusions, and disordered thought/behavior. However, people with schizophrenia also typically have impairments in basic cognitive processes, such as attention and working memory, and the degree of cognitive dysfunction is a better predictor of long-term outcome than is the severity of the psychotic symptoms.
Researchers have tried to identify the nature of cognitive dysfunction in schizophrenia since the 1960s, and our collaborative research group has spent almost 20 years on this problem. We now have a well-supported theory, which we call the hyperfocusing hypothesis, and we recently published a pair of papers that review this theory. The first paper describes the hyperfocusing hypothesis in detail and reviews the evidence for it, and the second paper contrasts it with the traditional idea that schizophrenia involves impaired filtering.
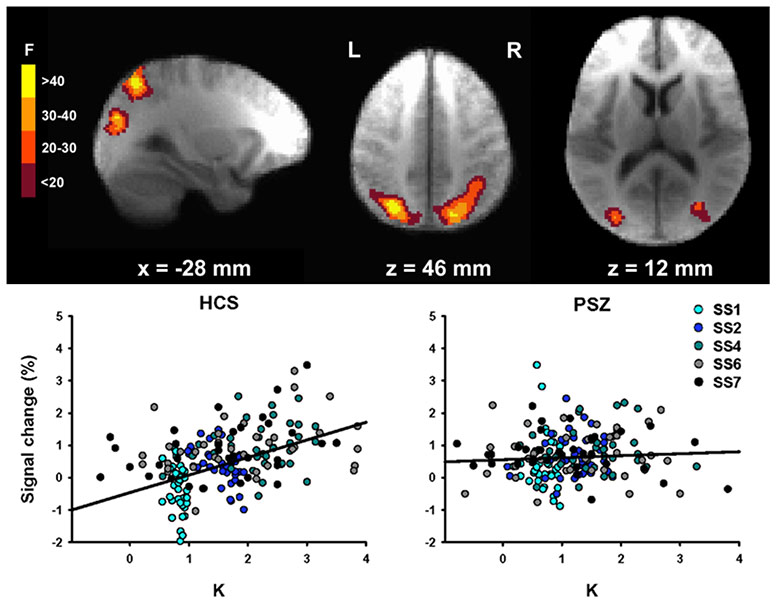
The hyperfocusing hypothesis proposes that schizophrenia involves an abnormally narrow but intense focusing of processing resources. That is, people with schizophrenia are not impaired at focusing their attention; on the contrary, they tend to focus their attention more intensely and more narrowly compared to healthy control subjects. This hypothesis can explain findings from several different cognitive domains, including reductions in working memory capacity (because people with schizophrenia have difficulty dividing resources among multiple memory representations), deficits in experimental paradigms that involve spreading attention broadly (such as the Useful Field of View task), and abnormal capture of attention by irrelevant stimuli that share features with active representations. In addition to explaining many previous findings, the hyperfocusing hypothesis has also led to many new predictions that have been tested and verified. We also find that the degree of hyperfocusing is often correlated with the degree of impairment in measures of broad cognitive function, which are known to be related to long-term outcome.
When a psychiatric group exhibits impaired performance relative to a control group, there are usually many possible explanations (e.g., reduced motivation, impaired task comprehension). However, the hyperfocusing hypothesis proposes that people with schizophrenia focus more strongly than control subjects, which leads to the counterintuitive prediction that people with schizophrenia will exhibit supranormal focusing of processing resources under some conditions. And this is exactly what we have found in several experiments. For example, in both ERP and fMRI studies, we have found that delay-period activity is enhanced in people with schizophrenia relative to control subjects when only a single object is being maintained. This is an example of what we mean by a “more intense” focusing of processing resources. You might be concerned that people with schizophrenia exert greater effort to achieve the same memory performance, and this leads to greater delay-period activity. However, when we examine subgroups that are matched on behavioral measures of working memory capacity, we still find that people with schizophrenia exhibit enhanced activity relative to control subjects when a single item is being remembered.
Classically, schizophrenia has been thought to involve an impairment in selective attention, a “broken filter.” For example, one individual wrote the following in an online forum: “Ever since I started having problems due to schizophrenia, my senses have been thrown out of whack... I remember one day when I got caught in the rain. Each drop felt like an electric shock and I found it hard to move because of how intense and painful the feeling was.” How can we reconcile this evidence for increased distraction with the idea that schizophrenia involves hyperfocusing? The most likely rapprochement between the hyperfocusing hypothesis and the broken filter hypothesis is that schizophrenia also involves impaired executive control, so people with schizophrenia often point their “spotlight” of attention in the wrong direction. As a result, they may focus narrowly and intensely on inputs that would ordinarily be ignored (e.g., drops of rain), producing greater distractibility even though the filtering mechanism itself is operating very intensely.